
Starvation induced down regulation of surface proteins
When cells are starved for nutrients, surface proteins are downregulated. Conserving energy from anabolic processes that are not essential during nutrient stress, and providing a catabolic supply of nutrients via lysosomal degradation. Some of our earlier research has shown some starvation responses to different nutrient depletion conditions:
Glucose starvation
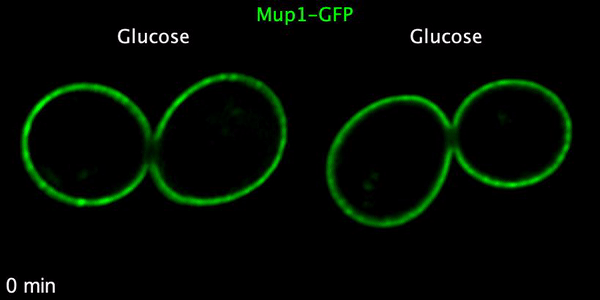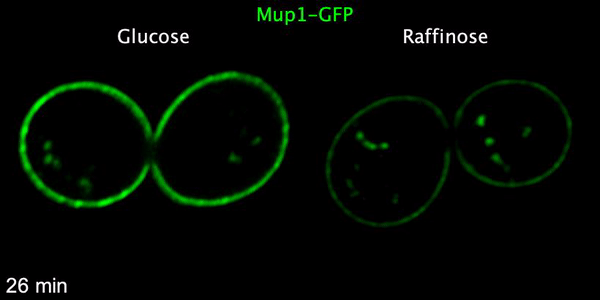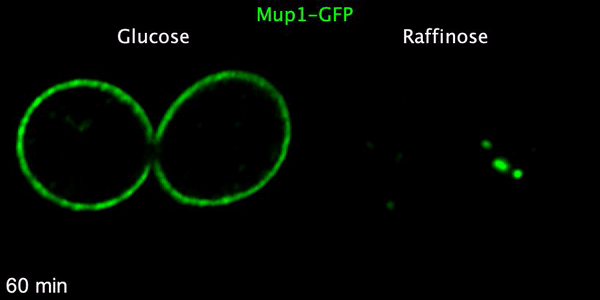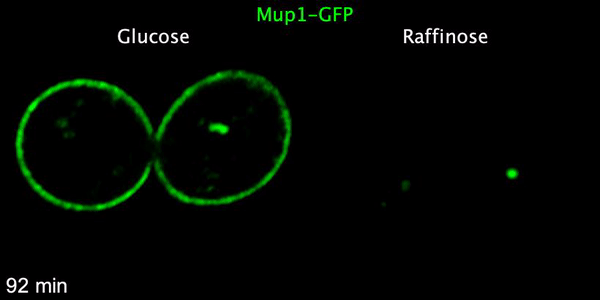
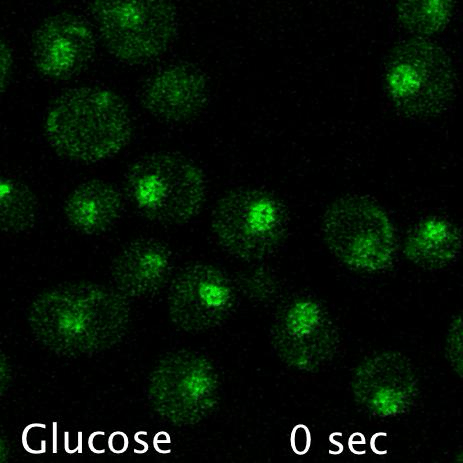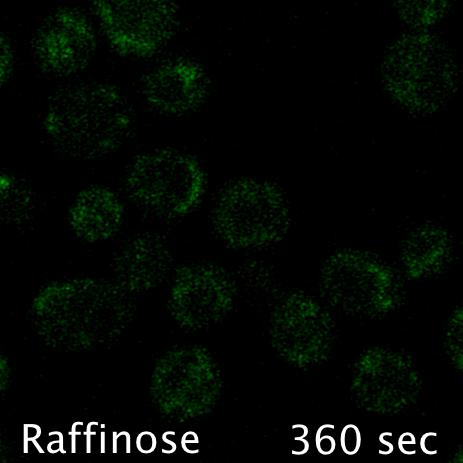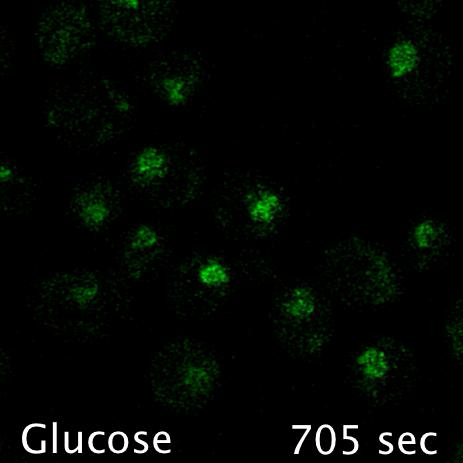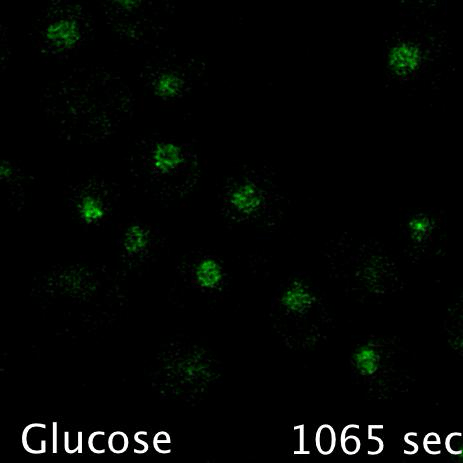
Using microfluidics to perfuse media with glucose (left) maintains GFP tagged surface cargoes (like Mup1) at the plasma membrane, whereas exchanging with raffinose (glucose starvation) triggers internalisation from the surface (right).
Mig1 controls down regulation of surface proteins
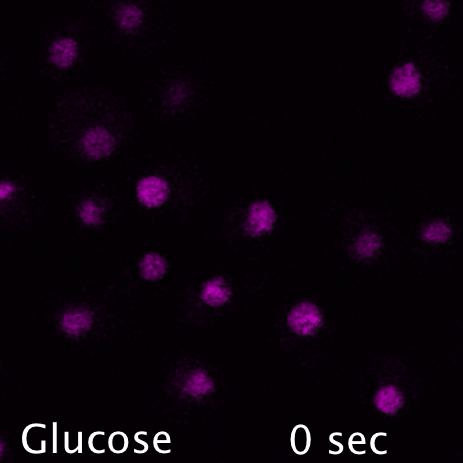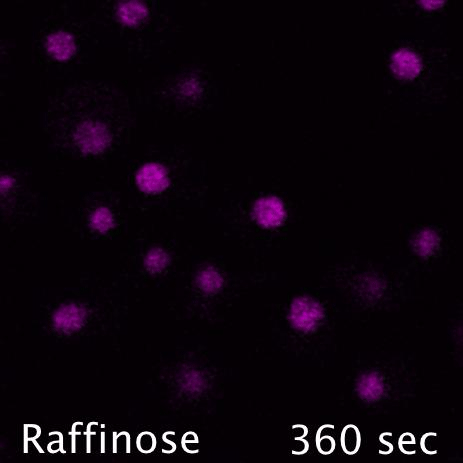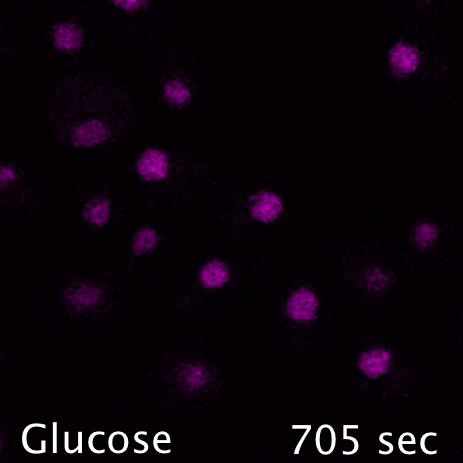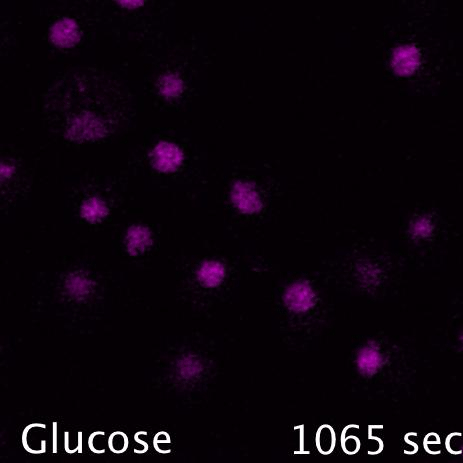
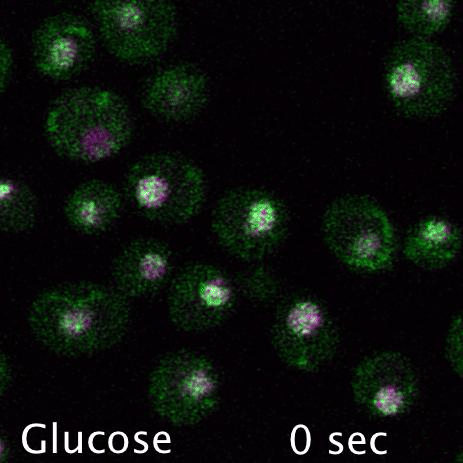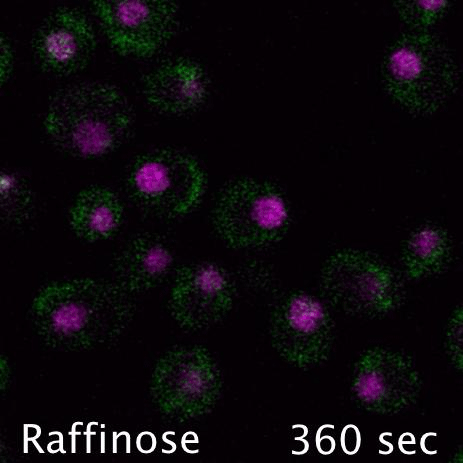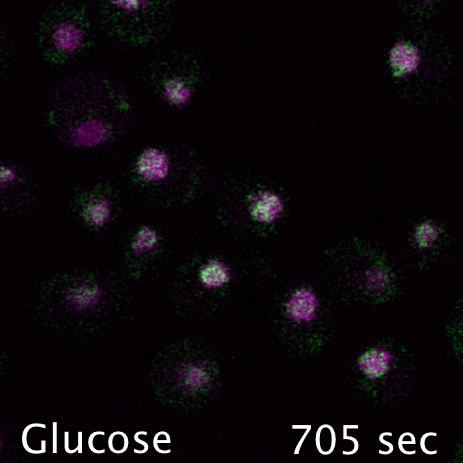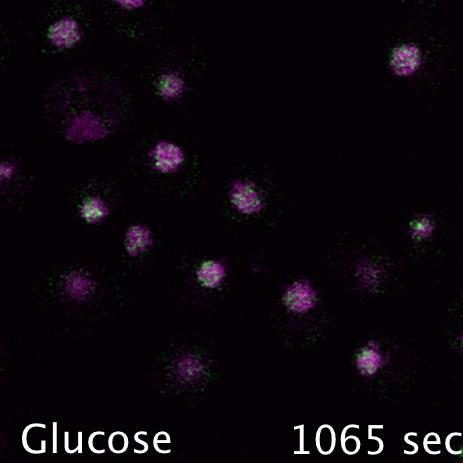
Mig1 (Green) is a transcriptional repressor that rapidly translocates from the nucleus (magenta) in response to glucose starvation. This triggers changes in membrane trafficking of two counteracting membrane trafficking pathways: clathrin mediated endocytosis and Ist1-mediated endosomal recycling.

Glucose starvation alleviates Mig1-mediated repression of YAP1801 and YAP1802, genes which encode the yeast AP180 clathrin adaptors (left). This increased in Yap1801/02 (right) is sufficient to downregulate surface cargoes.
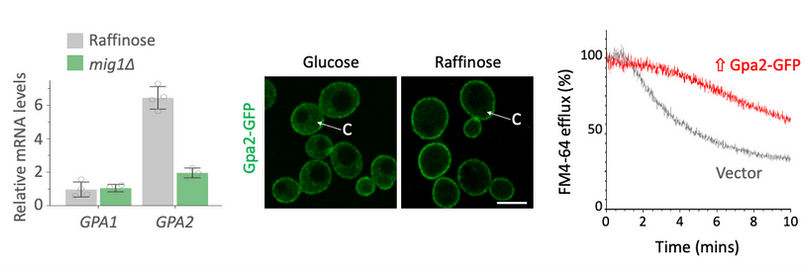
Glucose starvation also alleviates Mig1-mediated repression of GPA2 (left). Higher levels of Gpa1 in the cell, with increased concentration at the surface (middle) results in a decrease of endosomal recycling (right). The mechanism for this is Gpa2 physically interacts with the PI3K effector Gpa1, sequestering it at the surface and attenuating Gpa1-mediated recycling.
Eisosomes
In addition to down regulating surface proteins by increasing endocytosis and decreasing recycling, glucose starvation also spatiotemporally modulates surface proteins within the plasma membrane itself.
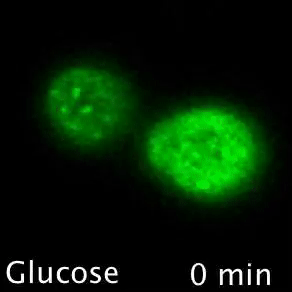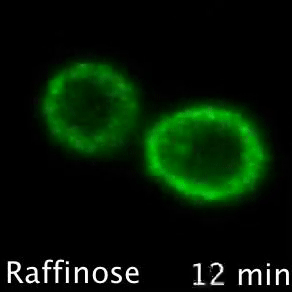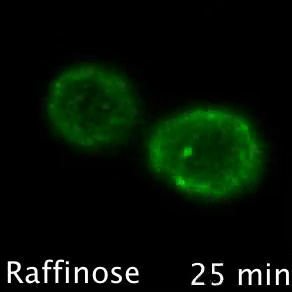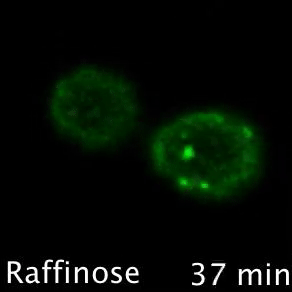
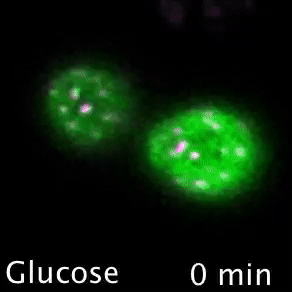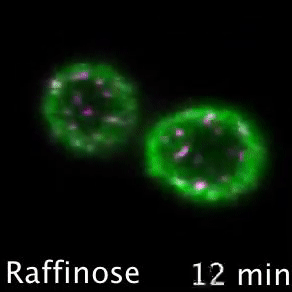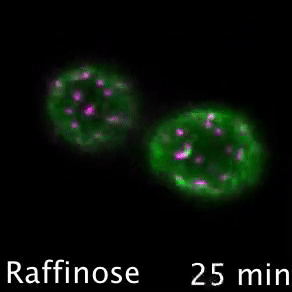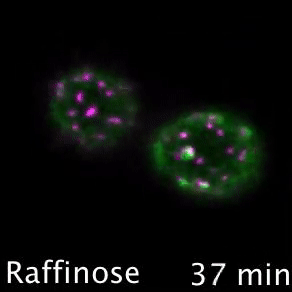
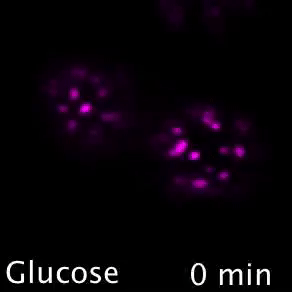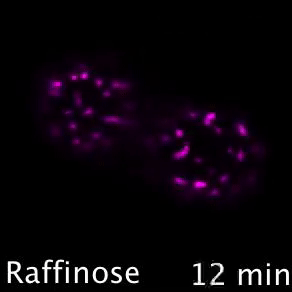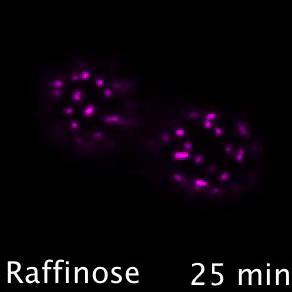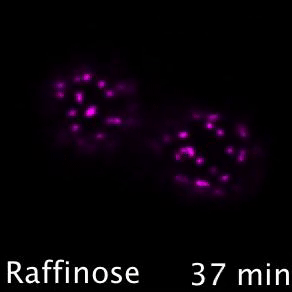
Nutrient transporters like Mup1 (Green) are largely degraded in response to starvation, but a small reserve pool is retained in eisosome subdomains (magenta) of the plasma membrane. This reserve pool is required for efficient recovery from starvation, with defects in eisosomes hampering recovery.
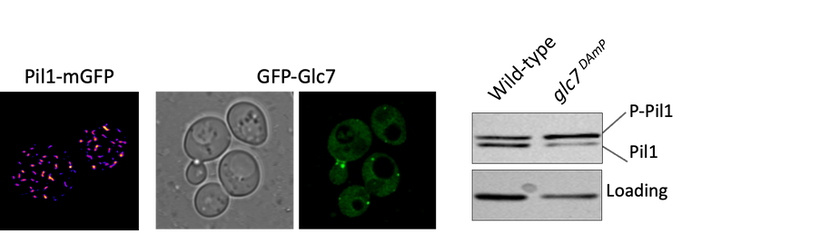
The BAR domain protein Pil1 is required for eissosome biogenesis (left). We performed an enzymatic screen to identify the phosphatase Glc7 (middle) that dephosphorylates Pil1 (right) in response to glucose starvation. Our model predicts that Pil1 membrane interactions - controlled via phosphorylation status - alter eissoomes to better harbour nutrient transporters during nutrient stress.
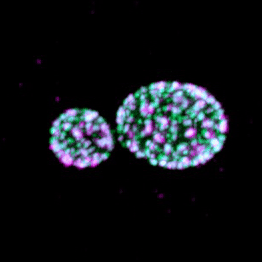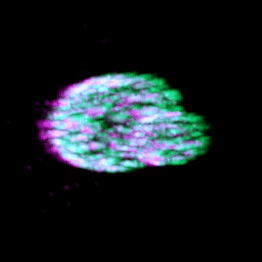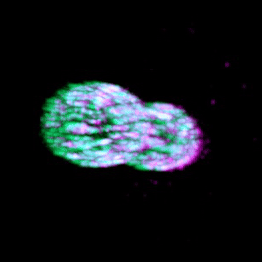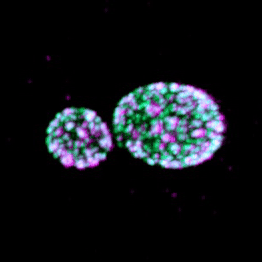
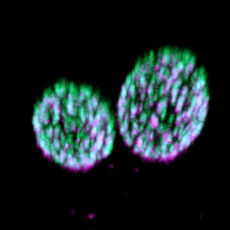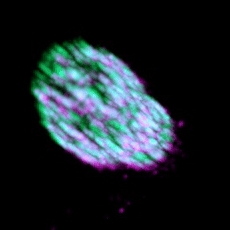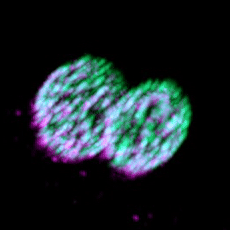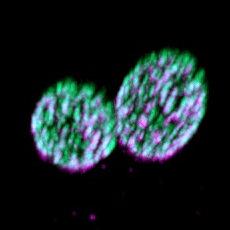
Eisosomes are not found in humans, but they are found in many different pathogenic fungi, including all four of the WHO's fungal priority pathogen list. Studying eisosome capacity to tolerate stress in such organisms as Candida albicans (shown) could lead to exciting theraputic potential.